
Genetics Of Immunity
Concept Of Immune Mechanisms
System of Immunity
Viruses, bacteria, parasites and several other pathogens surrdisound us at all times. Though some of these organisms reside normally in the body (commensals), some of them can breach the defense barricades and mechanisms of the body and cause diseases in humans. The body has, in fact, a well- developed system that fights such an attempt of invasion and thus prevents diseases that would perhaps be fatal had not the system been in existence.
The capability of an organism to prevent or modulate the occurrence of diseases both from within and without is called immunity. The system not only recognizes and destroys viruses, bacteria and other pathogens that attack the body from outside but also eliminates cancerous cells and toxins that arise within the body. The system is called the immune system for the function it serves.
The immune system has the unique ability to differentiate between cells and tissues that belong to the same body (self) and those derived from sources other than the same body (non-self) and decide selectively to preserve (self) or to attack and destroy (non-self) cells or tissues.
Read and Learn More Genetics in Dentistry Notes
Components of Immunity
Immune mechanisms operate chiefly through cells that circulate in the blood and mediate the actions of the immune system. The cells are the white blood cells called the lymphocytes. The formation of lymphocytes is initiated in bone marrow from the resident bone marrow precursor cells that are known as the stem cells. The undifferentiated cells gradually mature and differentiate into functional lymphocytes. The functional lymphocytes are of two different kinds, the T and B lymphocytes.
The T lymphocytes are differentiated and activated in thymus hence, also called as thymus-dependent cells. The B lymphocytes probably undergo differentiation as well as maturation in bone marrow itself and hence known as bone marrow derived cells. Lymphoid organs (lymph nodes, spleen, tonsils, Peyer’s patches, etc.) act as stores for most of the B cells in the body.
The T cells however are always actively circulating in blood stream. The other equally important component of the immune system is derived from the reticuloendothelial system called the macrophages.
Microorganisms and pathogens constantly attack our body and try to invade through wounds, discontinuous epithelial surfaces, the respiratory and the GI tracts, etc. and access the blood or any suitable place for them to multiply.
The immune system is activated in several steps by such an incursion by pathogens from outside or from developments within.
T-cell Immunity or Cell Mediated Immunity
A large number of T lymphocytes are activated that are involved in executing this type of immunity. T lymphocytes are specifically activated to identify signals on the pathogens and to destroy them. The action of these cells is also facilitated by the action of other immune cells. There are several types of T cells
(e.g. killer T cells, suppressor T cells and helper T cells) if the system detects even a single epitope on that cell that serve different functions.
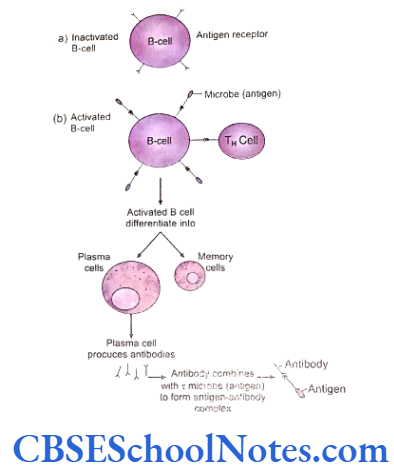
B-cell Immunity or Humoral Immunity
Humoral immunity is mediated by the B lympho- cytes. These lymphocytes are transformed to plasma cells under the influence of several factors. These plasma cells synthesize antibodies or immuno- globulins. Several varieties of immunoglobulins attack and destroy invading organisms.
Functioning Components of the B-cell Immunity
Antibody
Antibodies are defined as protein molecules synthesized by organisms in response to the presence of a foreign substance, in order to neutralize its effects. An antibody has a specific affinity for identification of and binding to the foreign material against which it is synthesized by a plasma cell. Antibodies are produced by plasma cells which are derived from activated and transformed B lymphocytes. A single plasma cell produces and secretes about 2000 identical copies of antibody molecules every second and secretion continues unabated for approximately 4 to 5 days till the plasma cell survives.
Antigen
Antigens are defined as immunity stimulating substances (a foreign macromolecule) that are capable of inducing antibody formation from B lymphocytes. Many substances are antigenic in nature (anything which is as big as or bigger than a protein molecule may act antigenic). Antibodies are produced against epitopes, the minimum sized protein molecules that can excite the formation of an antibody.
Basics of Immune Response
Specific and characteristic identifiable features are located on the surface of antigens like viruses, bacteria and cells of higher organisms that are called antigenic determinants (epitopes). A large molecule or cell may contain hundreds of different antigenic determinants or epitopes. The immune system in an individual gets activated against a cell or a substance inside the body that is different or absent in its own system.
A typical antigen includes several different epitopes and therefore induces the production of many different antibodies. Antibodies are produced in response to all varieties of antigenic determinants in an antigen.
B or T cells themselves are capable of detection of antigenic determinants of an antigen. They are facilitated by certain other immune cells for such an activity. Each B cell or T cell is capable of recognizing only one antigenic determinant on an antigen. Certain receptors on the surface of the T or B cells are configured exactly to match and bind with a specific antigenic determinant on the surface of an antigen (like a lock and key).
A determinant specific binding leads to cell division in the B lymphocyte. Mitosis is stimulated and the resultant daughter cells formed are of the same genetic constitution. These cells are called clones. Some of these clones differentiate to become antibody secreting plasma cells.
A particular clone is capable and destined to secrete just a single kind of antibody that is directed against the epitope of the antigen that initiated the B cell proliferation into a plasma cell. The subsequent combination of an antigen with its specific antibody is termed as antigen- antibody complex.
Complement System
The complement system comprises of about 20 plasma proteins that are triggered in a cascade on activation. This ‘activation’ is triggered with the formation of the antigen-antibody complex. The components of the complement system are activated sequentially and attack the antigen-antibody complex. The component of the complement system are basically certain activated plasma proteins capable of destroying trapped microorganisms and toxins in the system.
The complement system is a component of the immune system and hence acts either directly or indirectly (with activation of mast cells and macrophages) to destroy the offending pathogens in the body.
Diversity of Immune Response
There is a large diversity of antigens in the nature that interact with the immune system of our bodies’ day in and day out. As a consequence, there are a multitude of receptors on the surfaces of the immune T and B cells that bind to the different antigenic determinants. Since there are millions of antigens and antigens receptors, there are millions of varieties of antibodies.
It is a fact that antibodies are proteins and their synthesis is under genetic control. The obvious question comes up in mind that how a million of different antigen receptors as well as antibodies can possibly be generated from the available number of genes in the human genome.
To understand the mechanism of production of such enormous and diverse spectrum of antibodies we should first understand the structure of antibody (immunoglobulin).
Structure Of Immunoglobulins
Chemically immunoglobulins are glycoprotein in nature. Most antibodies contain four polypeptide chains. Two of these chains are long (each consists of about 450 amino acids) and are called heavy (H) polypeptide chains. These two chains are identical to each other. Short carbohydrate chains are attached to each heavy chain.
The two other chains are called light (L) chains or short polypeptide chains. They are also identical to each other and consist of approximately 220 amino acids.
Each light chain holds to the respective heavy chain with a disulfide bond. Two heavy chains are attached to each other approximately at their middle with the help of a disulfide bond. The region where the two heavy chains are connected is flexible and is called the hinge region. Because of this flexibility an antibody can assume a configuration that resembles the letter T or Y shape.
A heavy and light chain can be divided into two distinct regions. The tips of the H and L chains consist of the variable (V) region while the remaining region is called as constant (C) region.
The antigen binding region of the antibody is the variable (V) site. The antigen binding site of an antibody is typically very similar in structure to that of an epitope of an antigen to which it binds. The variable regions are different in each kind of antibody but are specific to the antigen it binds. The variable region is responsible for the detection and attachment to a particular epitope in a particular antigen. Most antibodies have two antigen binding sites.
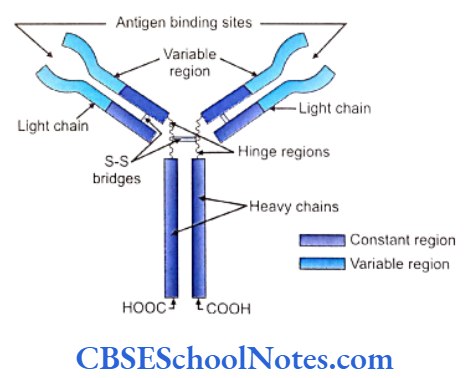
For all antibodies of a single class the constant (C) regions of H and L chains are nearly identical in structure.
The heavy chains are grouped into five classes of, Y, μ, α, δ and ε. The five different classes of immuno- globulins (antibodies) are determined by the chemical constitution of the five heavy chains. These different classes are IgG, IgA, IgM, IgD and IgE types.
Two different kinds of L chains exist in a given antibody namely the K (Kappa) or the λ (lambda) chains. Thus the molecular formula of IgG is δ2λ2 or δ2K2.
Approximately, one million antibodies are present in an individual and each of them differs in their antigen-binding specificity and affinity. It can thus be concluded that the variable region of an immuno- globulin molecule shows a wide range of variability; each with a different configuration. This structural variability is brought about by different combination of arrangements in the amino acid sequences at the variable end of the antibodies.
Determination of Diversity in an Antibody
- Chromosome number 2 and 22 bear the genes responsible for syntheses of the x and λ light chains respectively and a gene on the chromo- some 14 codes for heavy chains.
- The amino terminal ends (variable regions) of both the heavy and the light chains contain amino acid sequences of about 115 amino acids. Different types of antibodies constitute different sequences in the variable regions of both the heavy and the light chains. The carboxyl terminal end is made up of about 110 amino acids in the light chains kappa and lambda and forms the constant region C. The heavy chain has a constant region that is three to four times longer than that of the light chain.
- The DNA segments coding for the V region are separate from those that code for the C regions. This fact is established from restriction map study of the DNA segments that are responsible for coding of the C and V regions of K or light chains. The joining (J) regions that join the variable (V) and constant (C) portions of the antibody molecules are coded by the intermediate portions of DNA segments between the V and C coding regions.
- Placed between the V and J regions, the heavy chain possesses even a fourth region known as the diversity or D region (Fig. 7.3). Noncoding DNA sequences separate each coding region in the DNA segments (V, D, J and C coding sequences).
- Further, the variable regions of a given chain are coded by a large number of DNA segments. The D, J and C regions of the chain, in comparison, are coded by relatively few number of DNA segments.

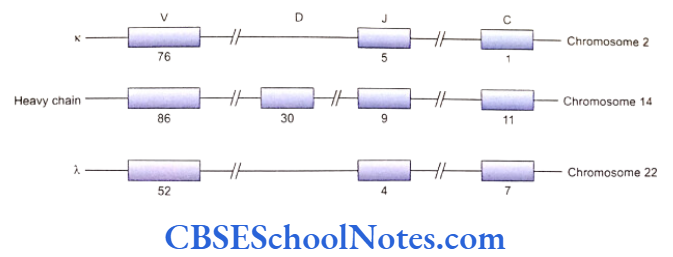
- A particular antibody molecule is assembled out of specific light and heavy chains. Each region (V, D, J and C) of a chain is again formed by unique sequencing of amino acids.
For example, the variable region (V) of a heavy chain is synthesized from one out of 86 possible genes coding for the variable region; the D region is expressed from 1 out of 30 genes; 1 out of 9 genes form the J region and 1 out of 11 shape the C region. Thus a single heavy chain is formed by 4 different genes taken from different available options. This recombination of several genes available for expressing different regions in the immunoglobulin imparts the diversity in the number and types of antibodies produced by the immune system. - Any specific variable (V) region gene of a heavy chain can be spliced on to any one of the (D) region genes. This combination can further be spliced on to any J region gene. This splicing process is called V-D-J joining that gives the genetic representation of the V-D-J segment of the antibody heavy chain. The constant portion gene of heavy chains (obtained from any one type of gene out of Cu, C8, and Ca, etc.) is now attached to the V-D-J segment to complete the heavy chain genetic sequence.
- After the splicing of V, D, J and C genes is over, it is followed by transcription. Transcription is followed by RNA processing. All the intervening sequences are removed during RNA processing.
The processed mRNA represents a gene containing all the four adjacent coding regions (V, D, J and C) of the heavy chain. This messenger RNA in a given B cell will thus produce a heavy chain with a specific variable (VDJ) and constant (C) regions, i.e. a specific type of antibody. All the progeny or clones of the same plasma cell will continue to produce identical antibodies.
Variability in antibody types results from:
- The possible combinations between large numbers of available genes that code for variable regions of heavy as well as light chains.
- Splicing of genes can also create altered codons at splice junctions. These codons generate new configurations in the molecule and hence an additional source of variation.
- Somatic mutation of antibody producing genes.
Immunodeficiency Disorders
Individuals are said to become immunodeficient when they exhibit lack of function of the cells of the immune system. The T cells, B cells or both may show a decline in their counts or effectiveness. Patients with T cell dysfunction suffer from viral illness but can accept mismatched skin transplants.
Lack of B-cell function enhances susceptibility to bacterial infections. These individuals may accept mismatched blood trans- fusions due to the absence of proper humoral immunity. Disorders of immunity may be inherited as genetic diseases or may be acquired from outside. A few of these diseases are summarized below.
Severe Combined Immunodeficiency (Swiss-type Autosomal Recessive Agammaglobulinemia)
Cellular and humoral immunities both are severely affected. Individuals are highly susceptible to both viral and bacterial infections.
Characteristics of the Disease
The disease has been identified as an autosomal recessive disorder. The absence or ineffectiveness of the enzyme adenosine deaminase is implicated in the disease. Mutation occurring in some T cell receptors and certain other T cell related proteins are responsible for syndromes similar to SCIDS. Affected children also suffer from the deficiency of granulocytes. Patients typically have low IgA and IgM levels in the serum. The thymus is usually absent or reduced in size in such cases.
Antibiotics are helpful in combating infections whereas bone marrow transplantation revives the WBC population.
Thymic Agenesis (Di George Syndrome)
Characteristics of the Disease
The absence of the thymus gland and subsequent maldevelopment of T cells result in this syndrome. Abnormalities in the development of the 3rd and the 4th pharyngeal pouches bring about the absence of thymus and also are associated with nondevelopment of the parathyroid gland. The disease is distinguished by severely depleted levels or absence of T lymphocytes.
Diseased children suffer from recurrent viral infections. Certain congenital heart diseases and tetany are also encountered in the syndrome.
Deletion of a particular region of the long arm of chromosome 22 is associated with the disease. Transplantation of fetal thymus in the patient may help prolong life.
X-linked severe combined immunodeficiency (swiss type X-linked agammaglobulinemia) and acquired immunodeficiency syndromes are examples of immunodeficient states.
Transplantation Of Tissues
It is seen that our immune system does not react against self antigens but mounts a severe response against non-self or ‘foreign’ antigens when they are introduced or transplanted into the host body. This unique character of the immune system to reject transplanted ‘foreign’ tissues and antigens is achieved by priming the immune cells to various self antigens during the early period of development of the fetus.
Cells reacting against the one’s own antigens during T and B cells that do not express receptors against the fetal life are discarded automatically by cellular events. ‘self’ and hence do not produce reactions (antibodies) against one’s own tissues are preserved for action in Thus when any tissue or organ from a person (donor) future against ‘non-self’ entities including pathogens. Is transplanted to an unrelated recipient, the lymphocytes of the recipient immediately recognizes the foreign tissue or organ.
These tissues act as an assembly of non-self antigens. Immune responses are mobilized against the transplanted tissue and the tissue is rejected. Transplants between identical twins and transplantation of tissues from one part of an individual to any other part of the same person can be done without rejection as the cells and tissues bear identical antigens that behave as self antigens and do not evoke immune responses.
Except for identical twins, tissue transplantation between individuals entails a detailed comparison between specific antigens between the individuals. Special tests predict the compatibility and success of such a transplant. The term histocompatibility denotes the evaluation of such a similarity between individuals.
Tissue typing is a process where the major and minor histocompatibility antigens (vide infra) of the donor and recipient tissues are examined and matched for their likeness (histocompatibility). Antigens on the donor cells, if not present on the cells of the recipient, results in the rejection of transplants.
Genetics of Histocompatibility
Specific proteins (antigens) called antigenic determinants are present on the surface of a cell. The expression of these antigens are genetically determined by alleles located at different loci (more than a dozen) known as histocompatibility loci. Each of the several loci produces a specific antigen, and each histocompatibility loci may contain one out of many available alleles. Therefore in whole of the human population each individual has a different combination of epitopes (antigenic determinants) on the cells due to unique combinations of alleles at those loci.
Thus an individual builds its unique genetic identity. As observed, stronger immune response is exerted by some of the histocompatibility loci and their alleles when compared to the others, i.e. some of the loci are more important than the others in context of HLA vide infra-compatibility. The major histocompatibility complex (MHC) loci are the most significant of all loci in the human race. There are of course many other minor loci in the genome.
In cases of transplantation between individuals incompatible for minor histocompatibility loci, some of the undesirable effects of transplantation can be treated with immuno- suppressive drugs. As these expressed antigens were first observed on white blood cells, human MHC locus is also called HLA (human leuokyte antigen) and it is situated on chromosome 6; the minor loci being scattered throughout the genome.
The HLA-A, HLA-B, HLA-C and HLA-D are the four different regions on the chromosome 6 that comprise the HLA loci. Each region of a locus again may be composed out of an ‘allele’ selected out of a series of available alternative “alleles”.
One allele for each of the A, B, C and D regions is present on each of the 6th chromosome. A haplotype is the make-up of the HLA alleles carried on each of the two chromosomes 6 in an individual. As obvious, different combinations of the HLA alleles on both the chromosome 6 provide innumerable possible HLA genotypes.
It is because of this that two unrelated individuals (belonging to different families, clans, countries, races, etc.) show a range of differences in their HLA geno and phenotypes. As expected though, the HLA make-up in siblings and in closer relatives are relatively similar to each other antigenically at their HLA loci. HLA constitutions in monozygotic or identical twins are exactly the same.
It is mandatory to conduct HLA typing tests in the donor and the recipient before any tissue transplantation is contemplated. The test is usually done by using a PCR based technique.
HLA and Disease
Curiously enough it has been found that the occurrence of as well as susceptibility to certain diseases are closely linked to the presence of a particular HLA type in an individual. An explanation for this might be a very close association or proximity of the disease producing gene to a particular HLA complex that segregate together during meiosis (gamete formation).
Genetics Of Immunity Summary
- The ability to resist the invasion of pathogens is brought about by the immune system.
- T-cell (Cell mediated) and B-cell (Humoral) immunity are the two subsets of the immune system.
- Cell mediated immunity involves several T-cells (killer, suppressor and helper) capable of destroying invading microorganisms.
- Humoral immunity involves B lymphocytes that are transformed to plasma cells which produces antibodies (immunoglobulins).
- Antigens are foreign macromolecules which are capable of inducing antibody formation.
- Antigenic determinants (epitopes) are identi- fiable features on the surface of antigens which can be recognized by B or T cell. Antibodies are always formed against an epitope.
- Antibodies are made up of four polypeptide chains. Two of the chains are heavy (H) and the other two are light (L). Disulfide bonds connect these chains with each other.
- The variable (V) region of the antibody is its antigen binding site. Variable (V) regions are different yet specific for each kind of antibody and thus can identify and attach to a particular antigen.
- Five different classes of antibodies (immuno- globulins) exist; IgG, IgA, IgM, IgD and IgE.
- Both the heavy and the light chains differ in their amino acid sequences in the variable regions. These sequences are different in each type of antibody. Thus a perfect antibody can be synthesized for a particular type of antigen.
- The variability in the structure of an antibody is achieved through different possible arrange- ment in the genes that code for the regions in the chains.
- Antigenic determinants (epitopes) of a cell or a substance are present on its surface. Before tissue or organ transplantation, similarities in certain antigens are assessed between the donor and the recipient. This is called histocompatibility testing.
- Antigenic determinants of great importance are expressed by genes located at histocompatibility loci (for both major and minor antigens).
- The composition of genes at the histocompatibility loci in an individual is unique due to specific combination of alleles at them. Thus each human carries a particular haplotype.
- The most significant histocompatibility loci in humans located on chromosome 6 is called the Major Histocompatibility Complex (MHC). The matching at the MHC determines the outcome of a transplant.