
Molecular Control Of Development
It is amazing to realize the degree of precision and detailing involved in the process of the development of a full-grown individual starting from the first cell of life, the zygote. The intricate mechanisms that control each of the steps in the process are nothing short of miracles of molecular engineering given the thousands of stages at which the process can veer towards undesired destination.
Thus though it may seem normal it is actually a matter of chance, so marvelously sustained as the usual’, that majority of the population walks around in normal physical and mental formats. The following sections of the chapter discuss the molecular mechanisms that guide and control the various processes of growth and differentiation in the developing embryo.
As detailing of each of the steps in molecular control of development is beyond the scope of this book, the description of the events would nevertheless enable the students of dentistry to understand the fundamental concepts of developmental genetics.
Embryonic development in humans or for that matter any kind of regulation imposed on the functioning of a cell, tissue or organ is enforced by expression of definite protein molecules. Eventually these are the genes that exert their influence on cellular functions by synthesis of a specific proteins needed for a particular function. The synthesis of proteins differs from cell-to-cell and within the same cell at different points of time. This provides the basic mechanism for control of any cellular process.
Read and Learn More Genetics in Dentistry Notes
The process of Growth in an organism is achieved by cell division through mitosis that multiplies the number of existing cells and also by increasing the amount intercellular matrix. The process of differentiation is the creation of new types of cells or tissues, which were not previously present. The differentiated cell possesses new morphological and functional characteristics, which distinguish it from other cells.
As described below, we now know that these characteristics result from the formation of new enzymes and proteins. Earlier workers tried to study the mechanism of differentiation by experiments on embryos of amphibia and chicks. Their work has produced many interesting results some of which are as follows:
It has been observed that certain regions of the embryo have the ability to influence the differentiation of neighboring regions. Interesting experiments have shown that these areas can induce formation of the same and specific tissues if they are implanted at areas outside their normal site of occurrences.
The primary organizer (identified at the dorsal lip of the blastopore) is the first organizer that is recognizable in the embryo. The failure of development of the primary organizer results in absolute failure of embryonic development. On the other hand if the dorsal lip of the blastopore is grafted on to a different site of another embryo, it induces the development of an entire embryo at the implanted site. Thus the signals that determine the initial organization in an embryo come from the primary organizer.
The effects of these organizers are brought about by enzymes or signaling proteins that are basically the product of gene transcription and translation. These signals may be in the form of (a) inductors which stimulate the tissue to differentiate in a particular manner; or (b) inhibitors which have a restraining There are stretches of DNA sequences called enhancers influence on differentiation.
Therefore the study of the controlling mechanisms can be termed Genetic control of development or described as Molecular control of development as it is now well-documented that the final control of the mechanisms of control rests with the genes involved in such control.
Though all the cells in the body contain the same complement of genes and other nuclear molecules, specialization of the structure and functions of a cell is determined by activation of only a certain number of genes in a particular type of cell. The process of protein synthesis involves two of the fundamental processes in cell biology; transcription and translation.
The basic process of transfer of genetic information begins with transcription of the mRNA molecule from DNA that occurs inside the nucleus followed by extrusion of mRNA outside the nucleus.
The sequential arrangement of codons on the mRNA is used for synthesis of proteins by translation occurring in the cytoplasm and involves protein synthesizing machinery in the cytoplasm, e.g. ribosomes, tRNA, etc.
In any given cell, at any given time (interphase), only a few of its genes are active and others are resting. Cells are said to be differentiated structurally and functionally because of expression of a small number of developmental regulatory genes (master genes) in them during specific time of embryonic development. The expression of such master genes initiate cascade of events in different cells imparting the cell and subsequently the tissues of the embryo their unique structural and functional identity.
Every differentiated cell contains two types of genes; the housekeeping genes and the specialty genes. The majority of genes (80-90%) in a cell are housekeeping genes which are required for basic cellular metabolic functions. These common genes are also widely expressed in other cell types of the body. The specialty genes are expressed to define the unique features of different cell types.
In higher organisms regulation of gene expression is quite complex and is brought about by the action of specialized molecules such as hormones or growth factors on the target cells. Regulation (either facilitation or suppression) of gene expression is effected through binding of a transcription factor to specific DNA segments in the promoter region of a gene (vide infra).
There are stretches of DNA sequences called enhancers that may be located within the noncoding sequences of the gene, or located upstream or downstream of the gene. These regions on the DNA can bind transcription factors and increase the rate of transcription. There are also similar regions which inhibit transcription and are called silencers. Transcription factors may bind to these specific regions on the DNA activating or inhibiting (turn on or turn off) gene expression.
Molecular Processes In Development
It is well known now that several genes and gene families play important role in the development of the embryo. Most of these genes produce transcription factors which control RNA transcription from the DNA template in the target cells. The transcription factors thus play an important role in gene expression as it can switch specific genes on and off by activating or repressing it.
It is believed that several transcription factors control gene expression, which in turn, regulates the fundamental embryological processes like induction, segmentation, migration, differentiation and apoptosis (programmed cell death) in embryonic cells till permanent cell lines are established in tissues. The above fundamental embryological processes are mediated by growth and differentiation factors, growth factor receptors and various cytoplasmic proteins.
Our existing knowledge regarding the molecular basis for embryonic development is mainly based on the Drosophila (fruit fly). However, evidences are now being gathered indicating that the basic body plan of mammalian embryo is under the control of many such similar genes as those in the fly that have been identified for controlling morphogenesis in Drosophila.
At the molecular level signaling is effected by protein molecules that act from outside the cells and can act locally or from a distance as intercellular signaling molecules. Many signaling molecules are called growth factors. Signaling molecules need to bind to receptor molecules that usually exist as trans- membrane proteins in the plasma membrane of the cells. Attachment of the signaling molecules to the receptor molecules cascade a series of events through which a molecular signal is relayed from the cell membrane to nucleus (signal transduction pathway) using related molecules.
Signal transduction is used by the cell to activate several mechanisms including generation of transcription factors which initiates gene expression in the nucleus. factors binds to the DNA at promoter or enhancer region of the specific gene and initiate the process of transcription. The transcription factors are important molecules that guide embryological development.
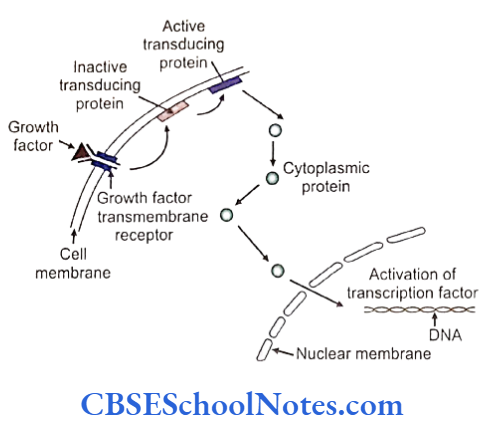
It is expected that mutations and disturbed expression of genes related to growth factors, receptors, or the transcription factors would be associated with various kinds of growth anomalies and cancers. The details of this phenomenon are discussed in appropriate sections of the book.
Growth and Differentiation Signaling Molecules
Proteins capable of stimulating cellular proliferation and cellular differentiation occur naturally and are termed growth factors. The epidermal growth factor (EGF), fibroblast growth factor (FGF) and the platelet derived growth factor (PDGF), stimulate the proliferation of epidermal cells, fibroblasts and the connective tissues, etc.
Growth factors typically act between cells in embryos through attachment to specific cell membrane receptors as intercellular signaling molecules. Several methods are adopted for execution of the effects of the signaling molecules. Growth factors can modify the expression or the effects of one another.
The signals are called hormones, which travel through blood to reach a distant place in the body. This system constitutes the Endocrine system.
Paracrine system act by signaling targets cells situated in the near vicinity of the signal executing cell.
The Juxtacrine mode of action requires that the effecter as well as the effected cells remain in cell-to-cell physical contact. The “gap junction” and “notch signaling are well known examples of juxtacrine system model of signaling. The notch signaling is described later in this chapter.
A few common growth and differentiation factor groups and their role in development are described below.
Growth Factors and their Functions
- Epidermal Growth Factor (EGF) Determines growth and proliferation of cells of ectodermal and mesodermal origin.
- Transforming Growth Factors (TGFs) TGF-B1 to TGF-B5 Forms the extracellular matrix, induces epithelial branching, myoblast proliferation.
Bone Morphogenetic Factors (BMP 1 to 9) Helps bone formation, cell division, cell migration and apoptosis.
Müllerian Inhibiting Factor (MIF) Regression of paramesonephric duct.
Nodal Formation of primitive streak, right- left axial fixation formation of mesoderm.
Lefty Determination of body asymmetry
Activin Proliferation of granulosa cells
Inhibin Inhibition of gonadotrophin
- Hedgehog proteins
Sonic Hedgehog, Desert, and Indian. Shh control neural tube formation, somite differentiation, gut formation, limb development, and growth of genital tubercle. - WNT Protein
Midbrain development, somite and urogenital ultimately activates a zinc finger transcription factor, Gli. differentiation, limb patterning. - Fibroblast Growth Factors (FGFs)
Mesoderm differentiation, angiogenesis, axon growth, limb development, development of various parts of brain, liver induction, mesenchymal proliferation in jaw, induction of prostate gland, outgrowth of genital tubercle., - Insulin-like Growth Factors (IGFs)
IGF-1 act as factor for bone growth, IGF-2 is a fetal chromosome number growth factor. - Nerve Growth Factor (NGFs)
Stimulate the growth of sensory and sympathetic neurons.
Abnormalities in the growth factor signaling pathway may lead to abnormal growth or cancer. The over expression of growth factors can lead to non- cancerous disorder like psoriasis. Mutation and over expression of PDGF gene may also cause cancers like osteosarcoma and astrocytoma. Mutation in growth factor receptors can lead to insulin-resistant diabetes (insulin receptor) and dwarfism (fibroblast growth factor receptor). Mutation and overexpression of these receptors are responsible for variety of cancers.
Growth Factor Receptors
Receptors are specialized protein molecules that recognize and bind specific signal molecules (ligands) such as growth factors and hormones. The transmembrane receptors are proteins situated across the plasma membrane of the cell. Receptors bind to the specific signaling molecules on the outer side of the membrane and activate certain molecules (G proteins, etc.) on the inner side of the membrane.
This is followed by a series of activation, mainly by phosphorylation, in some cytoplasmic proteins known as protein kinases, e.g. Tyrosine kinase, Protein kinase C, etc.
Other kinds of surface receptors also exist beside the transmembrane receptors. The notch receptor plays an important role in embryonic development. In this kind of signaling (juxtacrine signaling), a protein on one cell surface interacts with a receptor on an adjacent cell surface. Notch is a cell surface receptor, which has a long extracellular part and a smaller intracellular part.
Contact with the specific protein (delta or jagged) present on the surface of a nearby dominant cell activates the notch receptor. Attachment to one of these proteins causes the notch receptor to be broken in its intracytoplasmic domain. This broken portion acts as a transcription factor that regulates gene expression in that cell.
Thus one cell (called dominant) can influence transcription of an adjacent cell. Such example is visible in a developing neuron (dominant) that inhibits its surrounding cells to develop into glial cells. This phenomenon is termed as lateral inhibition.
Other growth factor receptors are responsive to molecules secreted by cells of the extracellular matrix (ECM) like glycoproteins, collagen, proteoglycans, etc. Receptors for fibronectin and laminin are called integrins.
For cell-to-cell communication, the gap junction channels are made up of connexin proteins.
Signal Transduction
The process by which a cell converts one kind of signal into another is called signal transduction. Binding of extracellular signaling molecules to receptors triggering a sequence of biochemical reactions inside the cell marks a signal transduction. These reactions are carried out by different enzymes as a chain of reactions and hence referred to as a “signal cascade”.
Extracellular growth factors regulate cell growth and differentiation by a complex pathway through signal transduction. Each sequential step in the pathway is genetically determined. The steps are sequenced as:
- Growth factor binding.
- Activation of the receptor.
- Activation of cytoplasmic proteins called signal transducing proteins; many of such proteins are situated on the inner surface of plasma membrane.
- Activation of one or more of the several cytoplasmic protein (kinase) systems.
- Formation of transcription factor.
- Effect of transcription factor: activation or inhibition the expression of a growth or a differentiation related gene.
We have already the idea that the processes in embryogenesis involve structures like ‘organizers’ that ‘induce’ the formation of specific types of cells from a common precursor. This process of induction is in fact carried out by organizers with the help of synthesis of several factors.
The synthesis of these important factors is under the control of certain well known genes that are involved with development of the embryo. One such gene is the PAX-6 gene which encodes for a transcription factor regulating several important events in embryogenesis including development of the eye.
Transcription Factors
A large number of transcription factors are common and found in all types of cells and across several organisms. However, few transcription factors are found only in certain types of cells or are active only during specific stages of development. The transcription factors regulate gene expression by acting on promoter or enhancer regions of specific genes.
These transcription factors are transcription regulatory protein molecules that bind to specific sites on the DNA. These proteins have ‘typical’ structural configurations at their binding sites with DNA. These sites are called ‘motifs’. Some of these ‘typical’ configurations are what we know as basic helix-loop- helix protein, zinc finger protein, etc.
Abnormalities in transcription pathways may lead to abnormal growth or cancer. The mutation of signal transducing proteins (e.g. ras gene) is responsible for almost 30% of human tumors. Mutations of genes which code for certain transcription factors are responsible for colon cancer, neuroblastoma, Burkitt’s lymphoma and lung cancer.
Specific abnormalities related to transcription factors are referred to in the section of the text dealing with molecular control of some important events in dental development.
Some of the important Transcription factors and their functions are discussed below:
- Basic helix-loop-helix protein is involved in myogenesis, neurogenesis, hematogenesis and the development of pancreas. This kind of transcription factors contain a short length of amino acids in which two alpha-helices are separated by an amino acid loop.
- Zinc Finger Proteins regulate expression of genes. The DNA binding domain in this protein is the zinc finger motif. The transcription protein is constituted by zinc ions binding to regularly placed cysteine and histidine units of the polypeptide chain. This results in puckering of the chain into finger-like structures (Fig. 8.2). These fingers configure to specific sites of the desired DNA helix. The kidney, gonads, hindbrain and white blood cells are some of the diverse examples of structures influenced by this transcription factor.
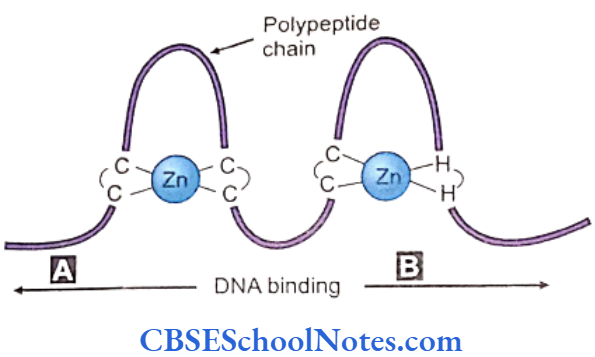
Anomalies in genes like GLI 3, WT1 and ZIC2 located in chromosomes 7p13, 11p13 and 13q32 respectively cause head, hand and foot abnormalities, Wilms’ tumor, ambiguous external genitalia and Holoprosencephaly. Mutations in the gene ZIC3 located at Xq26 may result in abnormal position of heart, liver and spleen.
- HOX genes regulate segmentation, patterning of the hind brain and formation of the axis of the embryo.
The HOX genes in humans encode a special class of transcription factors that regulate the sequential development of different body segments. Originally discovered in Drosophila this class of genes are called homeotic genes because mutations in these genes are capable of transforming one part of the body into another (e.g. growth of legs in place of antennae).
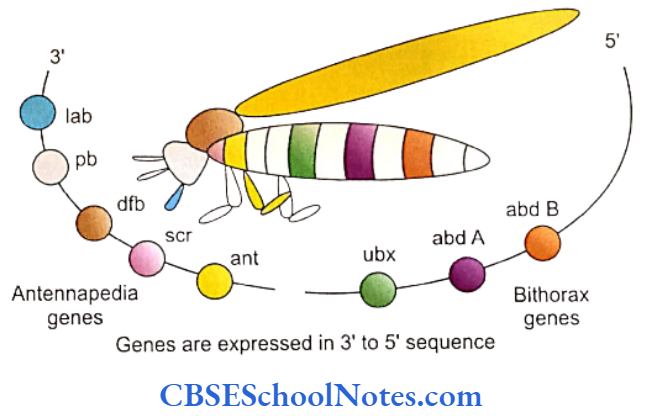
The regional morphogenic characterization of individual segments of Drosophila embryo is brought about by the expression of a group of homeotic genes. These genes determine which embryonic segment should bear antennae, wings or legs. These 8 homeotic genes are situated on chromosome number 3 and are arranged in two groups or clusters (Antennapedia and Bithorax). These genes are collectively called the homeotic complex or HOM-C.
Each of the 8 genes contains a highly conserved coding sequence of 180 base pair region of DNA (usually near their 3′ end) called the homeobox. The homeobox codes for a 60 amino acid protein called homeodomain. These homeodomains, thus, remain constitutively integrated within the bigger polypeptides coded by the homeotic genes.
These polypeptides synthesized by homeotic genes are transcription regulating factors. Homeo- domains within these polypeptides recognize and bind to specific DNA sequences of target genes.
The eight genes present in the homeotic complex express themselves in a selective sequence. That means the genes, which are cranial in position in the cluster or so as to say, located towards the 3′ end of the entire DNA material of the fly if put together in a 3′ to 5′ sequence, are expressed in the cranial segments or cranial regions of the developing embryo.
Successive downstream genes are expressed in the caudal structures of embryo giving each of the regions its structural identity in the craniocaudal axis. As stated before, the homeotic genes express polypeptide transcription factors. The products of a preceding or cranial gene regulate transcription in a succeeding or caudal homeotic gene.
Experimental mutation, suppression, or expression of the homeotic genes at different regions causes abnormal regional patterns in the embryo.
Such homeotic genes (clustered genes, each having a homeobox) similar to Drosophila are found in mammals and in humans. The human genes (called HOX genes) have same clustered organization, follow same order of gene arrangement within the cluster, their expression and functions are also in sequences as observed in Drosophila.
Quite interestingly, when the sequences within the homedomain and some short regions of the homeobox were compared, striking similarities were noticed across species. These sequences and their protein products (called ‘motifs’) have been strictly conserved through evolution. The amino acid sequences of homeodomains of Drosophila are up to 90% similar when compared with that of humans.
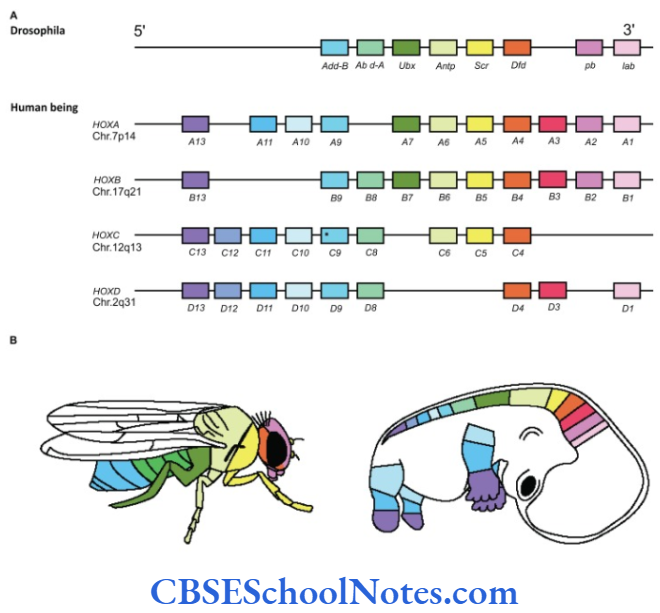
During hundreds of millions of years of evolution these genes have duplicated twice in man and hence human chromosomes have four copies of the clusters of homeobox genes. The genes (HOXA, HOXB, HOXC, and HOXD) arranged on four different chromosomes (Chromosome number 7, 17, 12 and 2). Genes in each group are numbered from 1 to 13 corresponding to the fly genes and each group can be placed in a vertical alignment.
Genes with same number but present on different chromosomes in the vertical alignment form a paralogous group. In humans, the HOXA and its paralogs are expressed in the cranial segments, the HOXB and its paralogs are expressed in the next caudal segment, the HOXC and its paralogs in the next segment and the pattern follows. The products of paralogs interact for a final result of expression within a segment.
There are 39 genes in all and each gene contains a homeobox region, which encodes for homeodomain protein. Similar to fruit fly, homeobox genes of humans are also expressed sequentially in craniocaudal direction during axis formation. The sequential expression of HOX genes correlates with the development of structures in craniocaudal sequence.
The HOX genes are responsible for cranial to caudal patterning of the derivatives of ectoderm, mesoderm and endoderm germ layers. HOX genes regulate the differentiation of somites, vertebrae and hindbrain segmentation. The expression of individual HOX gene may also occur in places like hair, blood cells and developing sperm cells.
This indicates that though the main function of HOX genes is to set up structures along the main axis of the embryo, but the individual gene may also guide the formation of specific structure, which may not lie along the body axis.
- PAX Genes (paired box genes) shape the development of sense organs (eye and ear) and the nervous system. These genes regulate cellular differentiation at the time of epithelial- mesenchymal transition.
The paired box gene is DNA sequence that encodes a 128 amino acid protein. This protein transcription regulating factor binds to the DNA at sites (domains) for activation of transcription. In humans the Pax gene family consists of 9 genes (Pax-1 to Pax-9). Details of the importance of these genes in dentistry is discussed later in the book.
Some of the developmental abnormalities associated with Pax genes are discussed below.
- The Pax-2 gene located on the 10th chromosome (10p25) when mutated results in renal malforma- tion and malformation of retine and optic nerve (renal-coloboma syndrome). Mutations in the Pax-3 (chromosome 2935), Pax- 6 (chromosome11p13) cause loss of hearing, areas of depigmentation in hair and skin and abnormal pigmentation of iris along with the absence of iris and sarcoma.
- SOX Genes (LEF-1, SRY type HMG) are expressed in many structures during develop- ment. Sox genes consist of over 20 members in the family. The Sox genes contain a 79 amino acid domain that is known as HMG (high mobility A BRIEF ACCOUNT OF THE MOLECULAR group) box. These genes show homology with Y-linked SRY gene. SRY gene plays a major role in male sex determination. The name of this group (SOX) was derived from SRY HMG box. The HMG domain activates transcription by bending DNA (hence, also called DNA bending protein) in such a way that other regulatory factors can also bind with promoter region of genes. The skeletal tissue and type II collagen development is linked to the Sox-9 gene. Mutation of Sox-9 on chromosome 17 result in bowing of long bones. A mutation of the Sox-10 on chromosome 22 is incriminated in Hirschsprung disease.
- POU Genes (Pit-1, Oct) Play a vital role in cleavage of the early embryonic cells. A Pou transcription factor is constituted by a homeodomain region and a second site on the factor that binds to the target DNA segment. The Pou gene family is named by the fist alphabets of few of the first genes identified i.e., Pit-1, Oct-1 and Unc-86.
The development of anterior pituitary gland is related to the expression of the Pit-1 gene. The Oct-2 gene is expressed in the B-cell activating immunoglo- bulin synthesizing genes. The Unc-86 gene is involved in the development of nematode neuronal cells. - Lim proteins regulate muscle differentiation. These genes constitute a large family and are associated with the development of all parts of body. Absence of Lim-1 protein results in headless embryo.
- T-BOX (TBX) Genes initiate the induction of mesoderm germ layer and specification of hind v/s forelimbs. Notochord differentiation is related to T-Box expression.
Also called Brachyury T-box genes, encode tran- scription factors that play important roles in develop- ment of mammary glands, upper limb and heart. Mutation of TBX-3 on chromosome 12 causes hypo- plasia of mammary gland and abnormalities in upper limbs. Mutation in TBX-5 may cause arterial septal defects and absence of forearm. - Dlx Genes (Dlx-1 to Dlx-7) are involved in morphogenesis of jaw and inner ear. The Dlx gene family consists of 6 members and is closely associated with HOX genes.
A Brief Account Of The Molecular Control Of Early Embryonic Development
In the preceding part of this chapter we have seen that the development of human body is regulated by sequential gene expression. In this process specific genes are expressed in sequence, one after the other, at different regions of the body resulting in development of dissimilar structures at different regions of the same body.
Finally the entire process of development results in structural and functional differentiation of highly specialized tissues and organs endowed with definite roles. These cascades of gene expression and the resultant sequence of embryological events are well-studied in the fly Drosophila that is very briefly discussed now.
Establishment of the Axes of Embryo
The process of axes differentiation and early embryonic development in Drosophila is one of the earliest events and under strict genetic control but in human this part of development occurs under lesser rigid genetic scrutiny.
In Drosophila the development of the anteroposterior, dorsoventral and right/left axes are under the control of a group of maternal genes, which are called maternal effect genes. These genes are expressed outside the egg (within the mother fly) even before fertilization. The products of expression of these genes are transcription factors called morphogens that subsequently act on future zygotic targets.
These products are carried into the egg where they diffuse unequally in the oocyte cytoplasm to establish gradients across the future anteroposterior axis of the egg. Due to the presence of such gradients proteins are differentially distributed in the common cytoplasm of the egg. These gradients determine the synthesis of specific proteins in the different segments of the embryo.
In the Drosophila body axes are established even before fertilization. In mammals body axes do not become fixed until the end of cleavage or early gastrulation. The formation of anteroposterior axis in human embryo is initiated by the cells of future anterior margin of the embryonic disk.
This area of disk expresses the genes (OTX2, LIM1, and HESX1) which are necessary for formation of the head even before gastrulation. The B-Catenin, BMP-4 and activin genes lead to the formation of primitive streak. These genes are first expressed in the cranial region of embryo. Once the primitive streak is formed the embryonic axes (craniocaudal, dorsoventral and right/left) are soon established.
Segmentation
The Drosophila embryo next divides into identical segments (Fig. 8.5). This is achieved by segmentation genes, which are subclass of genes called zygotic genes. The diffused ‘morphogens’ control the expression of segmentation genes. The segmentation is completed in three steps in the Drosophila embryo.
The segmentation gap genes control the first step of segmentation that divides the embryo into broad regions. Gap genes are controlled by maternal bicoid proteins.
The pair rule genes regulate the subdivision of the embryo in 7 segments along craniocaudal axis in the second step of segmentation. The pair rule genes are regulated by the products of genes regulating the previous step, the Gap genes.
Segmentation enters the third stage where the segment polarity genes like the Gooseberry, hedgehog, patched, wingless genes divide the embryo further into 14 segments. These segment polarity genes are controlled by the genes of the previous step, the pair rule genes. Similar segmentation genes used in developing humans have been identified.
Determination of Regional Characteristics
The process of segmentation is followed by development of regional characteristics in the newly formed segments of embryo. As discussed earlier, activation of the homeotic genes brings about specific characterization of individual segments of the developing embryo. These genes determine the growth of antennae, wings or legs in the appropriate segments of the fly.
The 8 Homeobox genes contains a highly conserved coding sequence of 180 base pair region of DNA (usually near their 3′ end) called the homeobox.
Similar to fruit fly, homeobox genes of humans are also expressed sequentially in craniocaudal direction during axis formation. The sequential expression of HOX genes correlates with the development of structures in craniocaudal sequence.
A number of other gene families that regulate development also contain similar homeobox domains synthesizing homeodomains but with different sequences in their genes, e.g. Paired, Pax (Pax-4 and Pax-6), POU, LIM, etc.
There is a direct relationship between vitamin A (retinol) and expression of HOX gene. Either too much or too little of retinoic acid (metabolite of Vitamin A) causes misexpression of HOXB-1. This may lead to abnormal development of legs, hindbrain and pharyngeal neural crest cells. The retinoic acid may cause extra pair of limb in frogs at the site of tail. This is an example of homeotic shift similar to formation of extra pair of wings in fruit fly.
Summary
- The process of Growth in an organism is achieved by cell division.
- The process of Differentiation is the creation of new types of cells or tissues.
- Thus the signals that determine the initial organization in an embryo come from the primary organizer.
- Molecular processes in development are governed by fundamental embryological processes like induction, segmentation, migration, differentiation and apoptosis (programmed cell death) in the embryonic cells.
- Cellular signals are relayed from the cell membrane to nucleus in sequential steps (signal transduction pathway) for the initiation as well as control of cellular processes.
- Several molecules are involved in the process of signal transduction like growth and differen- tiation signaling molecules, e.g. Epidermal Growth Factor (EGFS), Transforming Growth Factors (TGFs), Hedgehog proteins, the WNT proteins, etc.
- These Growth factors bind to their receptors to execute specific signal transduction activities.
- Transcription factors are the molecules that interact directly or indirectly with the genomic DNA to carry out final effects of cell signaling. Proteins like the Basic helix-loop-helix protein, Zinc Finger Proteins, HOX gene and Sox Gene proteins, etc. are important transcription factors.
- The process of early embryonic development includes the steps of establishment of the axes of embryo followed by segmentation of the embryo and determination of its regional characteristics.